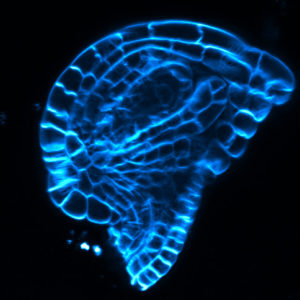
Research in the Shpak lab investigates the role that cell-cell communications play in development of aboveground plant organs. We are especially interested in the ERECTA family (ERf) signaling pathway. This pathway regulates multiple aspects of plant development. Currently, we are studying its involvement in the regulation of shoot apical meristem maintenance, leaf initiation, and flower development in Arabidopsis thaliana.
The ERECTA family signaling pathway
ERECTA family (ERf) receptor kinases are localized in the plasma membrane. Their extracellular domains, made of 20 leucine rich repeats (LRRs), bind small proteins from the EPF/EPFL family. The co-receptor TMM defines specificity of EPF/EPFL binding. Indirect evidence suggests that ligand binding leads to formation of heterodimers with SERKs, another family of receptors kinases, and to phosphorylation of downstream targets. A MAPK cascade functions downstream of ERfs; however, precisely how it is activated by ERfs is not clear. One of the targets of this cascade is the transcription factor (TF) SPCH. In Arabidopsis, there are three ERfs: ERECTA, ERL1, and ERL2, and eleven potential ligands.
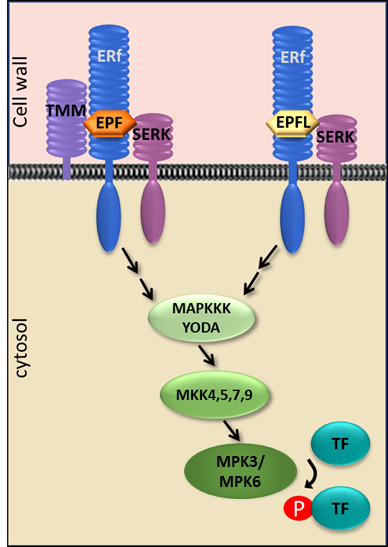
ERfs AND SHOOT APICAL MERISTEM MAINTENANCE
The shoot apical meristem (SAM) is a small but complex structure at the top of the stem that over time forms above ground organs: internodes, leaves, and flowers.

The SAM must balance the maintenance of stem cells in the center with differentiation of cells into organs at the periphery.
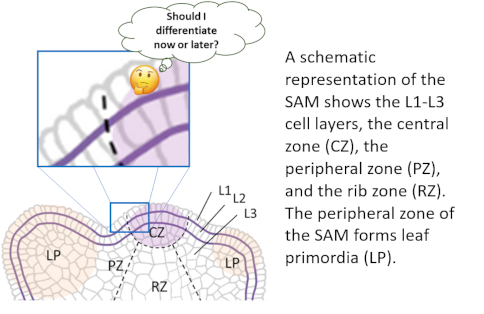
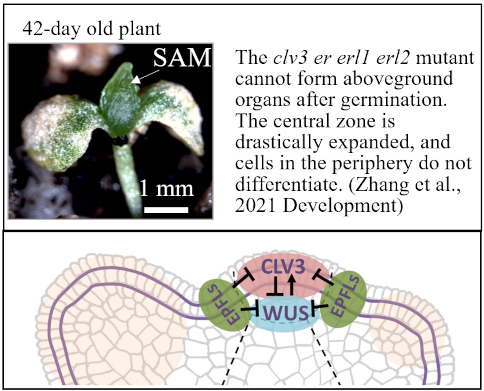
Synergistically with the CLV3 signaling pathway, ERfs restrict the size of the central zone and promote differentiation of organs. EPFLs are expressed at the periphery of the SAM and laterally restrict expression of the transcription factor WUS that promotes proliferation of stem cells in the central zone. EPFLs also inhibit expression of CLV3 and several other central zone genes.
Current projects in the lab
- The study of the molecular mechanism through which the ERf/EPFL signaling pathway restricts expression of CLV3 and WUS.
- The role ERfs play in regulation of other meristematic genes.
- In collaboration with Dr. Tian Hong’s lab, we are using mathematical modeling to understand patterning of the SAM.
ERfs PROMOTE INITIATION OF LEAVES AND REGULATE PHYLLOTAXY
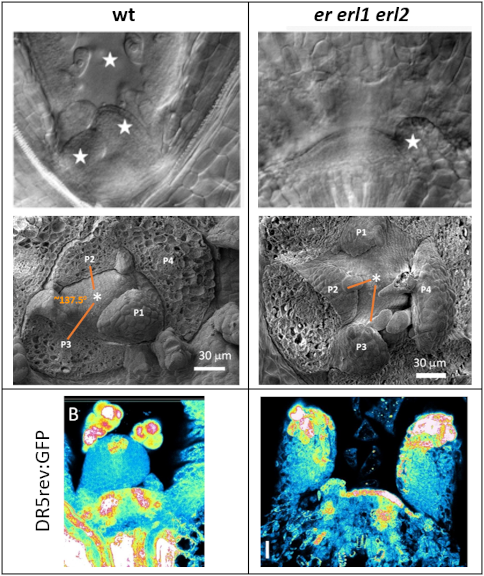
The central function of the SAM is to produce organs. In Arabidopsis, consecutive leaves and flowers are formed in the periphery of the SAM at 137.5 degree angles to each other, producing a spiral pattern around the stem. Their initiation occurs at a specific distance from the center of the SAM. Plant hormone auxin is essential for establishing the phyllotactic pattern.
ERfs promote leaf initiation and are critical players in establishment of phyllotaxy. The vegetative SAM of the er erl1 erl2 mutant initiates leaf primordia at a significantly reduced rate with almost random divergence angle. An analysis of auxin distribution using the DR5rev:GFP construct suggests that auxin distribution or responses are altered in the SAM of the er erl1 erl2 mutants. Currently, we are investigating the role ERfs play in modulation of auxin responses.
Cell-cell communications during flower differentiation
We study the function of ERf-EPFLs during the formation of a variety of flower structures including ovules, integuments, and anthers with the goal of identifying processes similarly controlled in different tissues. Specifically, we are testing the hypothesis that ERf-EPFL signaling controls morphogenesis by specifying boundaries between outgrowing structures. This research is funded by the National Science Foundation (NSF), https://grantome.com/grant/NSF/IOS-2016756.